How the end-Cretaceous mass extinction drove the evolution of modern snakes

The pitviper Gloydius intermedius, from the Gobi Desert of Mongolia. (Nick Longrich)
SNAKES are one of the most successful groups of tetrapods, which is a little ironic, because the key feature of the tetrapods (Tetrapoda is Greek for ‘four-footed’) isn’t even found in snakes. Despite this, or because of it, snakes are astonishingly diverse. With well over 4,000 living species, they’re the most species-rich group of reptiles. One might assume that the origins of this diversity are ancient, going all the way back to the snakes’ first appearance in the age of the dinosaurs, over 100 million years ago. In fact, modern snakes seem to have a far more recent origin. Fossils and DNA both suggest that modern snakes have radiated and become widespread only after the end-Cretaceous mass extinction that wiped out the dinosaurs. In the wake of the asteroid impact, snakes exploited niches and occupied habitats left vacant by an extinction that wiped out perhaps 90% of all species on Earth.
The Origins of Serpentine Diversity
Just how diverse are snakes? At last count, there are 3,921 species, yet about 20% of those have just been found in the last decade, and dozens are still found every year. That implies we’re nowhere close to finding them all.
Snakes aren’t just diverse in terms of species. They occupy a huge range of habitats— deserts, rainforest, grasslands, deep oceans. All snakes are predators, but they eat a huge range of prey- invertebrates like insects, snails, and centipedes, small vertebrates like fish, frogs, birds, and small mammals, and other snakes. A few snakes eat large mammals, sometimes humans.
In terms of locomotion, the seemingly simple snake body plan is capable of great flexibility (no pun intended) in terms of how it moves. Snakes can crawl, burrow, climb, swim; a few even fly.
The origins of snakes go back to the days of the dinosaurs, with the earliest definitive snake, Tetrapodophis amplectus, occurring in the Early Cretaceous Crato Formation of Brazil, around 115 million years ago. Tetrapodophis is far from a modern viper or garter snake however. It retains well-developed limbs, four of them. At this point, snakes don’t appear to have been either particularly diverse or widespread.

The oldest definitive snake is the four-legged Tetrapodophis amplectus from the Early Cretaceous of Brazil (Helmut Tischlinger)
At the start of the Late Cretaceous, fossil snakes become more diverse. In the Cenomanian of South America, we have the primitive snake Najash. Najash had lost its arms, but retained well-developed legs with hips, thighs, and knees. It isn’t closely related to any living snakes. Other primitive snakes are found about the same time in North Africa, including Lapparentophis, Norisophis, and Simoliophis. Simoliophis and its relatives (family Simoliophiidae) also retained legs (we don’t have good fossils of the others).
This diversification continued into the end of the Cretaceous. Large-bodied snakes, like Dinilysia from Argentina, Cerberophis from Montana, and Sanajeh from India appeared, along with the burrowing Coniophis and another aquatic lineage, Nigerophiidae.
The affinities of these Cretaceous species are still controversial, partly because the fossils are so incomplete. But where we do have good fossils (skulls and skeletons for Dinilysia, jaws for Coniophis) these are invariably archaic snakes, either distantly related to living snake families, or outside of modern snakes altogether.
Even by the end of the Cretaceous, few fossils can be linked to modern snakes— boas, pythons, garters, cobras, vipers, and so on. The modern snake families only appear in the fossil record later, millions or tens of millions of years after the dinosaurs went extinct.
Snake Extinction, and Radiation
What do these patterns mean? Well, the conventional wisdom long held that the extinction of the dinosaurs left many other groups, like amphibians and reptiles, relatively unaffected. It was just the big stuff going extinct, little things were largely spared. The snake fossil record suggests otherwise.
In fact, the snake record is similar to the bird fossil record. Here, the Cretaceous bird fossils are dominated by archaic species. The primitive birds all vanished at the end of the Cretaceous, and were then replaced by modern-looking forms- swifts, ducks, owls, penguins, tropicbirds, parrots, flamingos, etc., in the Cenozoic.
This appears to result from a massive extinction of archaic birds across the Cretaceous-Paleogene boundary, when the asteroid wiped out the dinosaurs— and perhaps 90% of all species on Earth. The extinction was then followed by a rapid radiation of modern birds in the aftermath.
So could the same be happening with snakes?
Maybe.

Cerberophis is a large (~2 meter) snake that lived alongside the last dinosaurs, including T. rex and Triceratops. Its known from just one vertebra- so it’s not entirely clear what it is- but the species disappears when the asteroid hit. Longrich et al. 2012 Longrich et al. 2012.
The problem is that the snake fossil record is incredibly incomplete. How bad is the snake fossil record? It’s so bad, even things like hominins and birds, which are notorious for having an incomplete fossil record, start to look pretty good compared to snakes.
What’s more, most of the fossils we do have consist of little more than isolated vertebrae, which are difficult to classify. Boas and pythons, for example, have very similar-looking vertebrae, but aren’t very closely related. Similarities in their vertebrae may reflect convergent evolution for a similar lifestyle, not necessarily close relationships. Between the scrappy fossils and the convergence, much of the Cretaceous stuff is just hard to classify. That makes it difficult to say for certain which lineages survived, and which went extinct.
A way to get around the issues of the fossil record is to look at another source of data on the evolution of Cretaceous snakes- the DNA of living snakes. The DNA of modern creatures is, after all, the product of tens and hundreds of millions of years of mutation, selection, speciation- if we could reconstruct the patterns of evolution, we might be able to make some sense of snake origins.
Using that DNA, for example, we can employ molecular divergence dating- otherwise known as a ‘molecular clock’.
The idea behind the molecular clock is that mutations gradually build up in a species’ DNA over time, like grains of sand in the bottom of an hourglass. If we can count the mutations in a lineage, and know something about the rate at which mutations appear, we can estimate how long it’s been since a species split off from its nearest relatives. It’s a little like one of those train problems in math class: “two snakes evolve at a rate of 10 mutations per million years, and 200 mutations separate them. How long ago did the two species separate?”
Using this technique, we can determine when various speciation events happened, at least, in theory. (As Yogi Berra famously put it, “In theory there’s no difference between theory and practice. In practice, there is.”)
The molecular clock has previously been used with birds. Although early attempts to model bird evolution suggested that most modern groups emerged at the end of the Cretaceous, more sophisticated approaches, with better constraints on key divergences, better genetic data, and more sophisticated models, suggest a far more recent origin. In this model, the genetic diversity of living birds is best explained by a pattern in which a handful of bird lineages- perhaps as few as three species- survived the asteroid impact that ended the Cretaceous, 66 million years ago, and then diversified, giving rise to all modern species.
Could snakes be doing something similar?
To test this hypothesis, my colleagues undertook a molecular clock analysis of snakes. Now, I’m going to assume that 99% of the world has no interest in the details of the assumptions and constraints and models that go into the molecular clock (If you’re part of the 1% (probably far less) who DO care about this stuff, feel free to read the SI of the paper. it’s complex, but in short I’d argue that (1) “extraordinary claims require extraordinary evidence”, that is, we should be highly skeptical of scrappy fossils that push lineages very far back in time, and that (2) both in theory and in practice(!) the most realistic model for a molecular clock is one called the autocorrelated clock, which assumes that related species tend to evolve at similar rates).
So. What did we find?
Remarkably, the vast majority of modern snake diversity is younger than the dinosaurs.

The evolutionary tree recovered by our analysis suggests that the ancestor of all modern snakes lived in the Cretaceous, ~100 million years ago- when snakes start to diversify in the fossil record. Yet only a handful of modern lineages extend back into the Cretaceous- perhaps five or six lineages.
The vast majority of all other snake lineages (again, your boas, pythons, cobras, garters, vipers, racers, sunbeam snakes, burrowing pythons etc. etc. etc.) diversify after 66 million years.
The fact that so few lineages extend into the Cretaceous is consistent with the idea that the asteroid impact at 66 MYA wiped out much of the Cretaceous snake diversity. Five or six survivors gave rise to everything alive today.
And number of fossil snakes in the latest Cretaceous- Dinilysia, Cerberophis- disappear and leave no descendants, suggesting there is some turnover here.
To be fair, an evolutionary tree of modern species can’t account for snakes- like Madtsoiidae, Coniophiidae, and Nigerophiidae- that survive the end-Cretaceous mass extinction, only to go extinct later in the Cenozoic. Clearly, more than just six species survived- although not all of those lineages make it today. They lucked out and survived the asteroid, but their luck ran out over the next 66 million years (a similar pattern is seen in some mammals- multituberculate mammals did pretty well surviving the asteroid impact, but ultimately were wiped out).
Still, the pattern is clear that not much crosses that boundary successfully. And the fact that so many lineages appear afterwards strongly argues that an adaptive radiation in the aftermath gave rise to modern snake diversity.
Again, that resembles the bird pattern: few things cross the Cretaceous-Paleogene boundary, lots emerge afterwards.

Time-calibrated DNA tree for modern birds, showing a handful of lineages extending before 66 MYA, and rapid appearance of lineages just after the asteroid impact at 66 MYA. (Prum et al., 2015)

Here’s the other half of the bird tree- so much happens pos-66 MYA it’s hard to fit on a single page.
And mammals (below) show a remarkably similar pattern. Cretaceous mammals suffered extremely high extinction rates, above 90%, and all modern placentals descend from perhaps 12 survivor species:

Time-calibrated DNA-based tree (molecular clock model) showing few boundary-crossers and multiple radiations of mammals after the end-Cretaceous mass extinction. (Dos Reis et al., 2012)
“And I Only am Escaped Alone to Tell Thee”
Yet while snakes were hit hard, some made it. And evolution is a game of relative success. Even if most snake species were wiped out, compared to ornithischian dinosaurs, plesiosaurs, pterosaurs, and ammonites- which all suffered 100% extinction- snakes were winners. Even 99% extinct is infinitely better than 100% extinct.
So why did (some) snakes make it when dinosaurs and so many other species didn’t?
Probably a number of factors contributed to snake survival. As Cretaceous snakes were fairly diverse, those factors weren’t necessarily the same for all species. But probably, the ability to burrow, to hunt in the dark, and to go long periods without food all helped, as did their prey preferences.
For one thing, with long, skinny bodies, snakes can find shelter in burrows underground.
Following the impact, the surface may not have been a safe place to be. Even though there’s little strong evidence for the more extreme ‘firestorm’ scenarios with widespread fires, it’s possible that the ejecta, shot into space and now re-entering around the earth, would have been hot enough to kill animals left out in the open. If so, burrows may have provided a shelter from the initial impact.

Wormlike blindsnakes- here, a little blindsnake I found on Zanzibar, possibly Indotyphlops brahminus- lived underground, eating bugs, where they could ride out the asteroid impact. Three different lineages of blindsnake survived the extinction. (Nick Longrich).
Afterwards, fine dust and soot shot into the high atmosphere would have blocked the sunlight from reaching the surface, causing temperatures to plunge. Again, burrowing underground would have offered protection against the cold until temperatures rose. Highly specialized fossorial snakes- blindsnakes- would have hardly noticed, since they spent almost their whole lives underground (note that a number of other subterranean lizards- at least two lineages of Amphisbaenia, and the Dibamidae- also survived the extinction).
Debris in the atmosphere would also have reduced the amount of light reaching the surface to the point that it became difficult to find food using vision. Snakes, which use nonvisual cues to find prey — flicking their chemoreceptive tongue, and sensing vibrations — may have had an advantage in a world of perpetual darkness (particularly for blindsnakes- which hunted in total darkness belowground anyway).
For carnivorous snakes, their feeding strategy and physiology probably helped them survive. In the cold and darkness, photosynthesis would have shut down, plants would have stopped growing, and animals would have begun to starve. Food was scarce.
But snakes can use a binge-and-fast feeding strategy, regularly eating animals as big as they are- and then going weeks and months before their next meal. A snake might emerge from its burrow and take a mammal, then retreat belowground for weeks or months, waiting out the worst of the extinction.
What’s more, being cold-blooded, their metabolisms would have slowed down when temperatures fell, making it easier to wait out the long, lean period of impact winter. Warm-blooded mammals, birds, and dinosaurs had to expend more energy to stay active in the cold- even as food became scarce. When it’s cold and food is hard to find, being warm-blooded becomes a liability.
What food was available, however? Darkness would have caused photosynthesis to collapse, leading to the death of plants, and the animals that ate those plants. Herbivores- animals like Triceratops and duckbill dinosaurs, but also herbivorous mammals and reptiles- would have starved to death and gone extinct. The same would have happened to their predators, animals like T. rex and velociraptors.
Some kind of food chain would have emerged, but it would have been based not on plants, but on scavenging dead stuff.
Dead leaves and trees would have provided food for invertebrates like termites, beetle larvae, and earthworms. For insect-eating snakes- the Typhlopidae, Leptotyphlopidae, and Anomalepididae- all of which cross the Cretaceous-Paleogene boundary- the specialization on termites, ants, and other soil-dwelling invertebrates might have meant that they had a glut of food when everything else was starving.
Dead dinosaurs would have fed beetles and flies, and the mammals gnawing their bones. Fungi and mold overgrew everything, feeding more insects. These invertebrates would have fed other predators- lizards, frogs, birds, mammals. And of course, snakes could have eaten those predators. It wasn’t much of a food chain, feasting on those who feasted on the dead, but it was something.
Meanwhile, the rain washed debris- leaves, dead trees, dead dinosaurs- into lakes, streams, rivers, estuaries. This dead matter fed invertebrates like insects, worms, snails, which then fed small vertebrates like fish, frogs- then larger vertebrates, things like alligators, turtles, and snakes. This is thought to be why animals like alligators, turtles, salamanders, paddlefish, and so on did fairly well during the extinction (that, and water has extraordinarily high thermal inertia, protecting from both heat, and cold).
At least one lineage of snakes that survived the Cretaceous- the now-extinct Nigerophiidae- were aquatic, and they likely exploited freshwater ecosystems for food, and may have found shelter there against extreme temperatures.
“The winds and waves are always on the side of the ablest navigators.”
The success of modern snakes isn’t due to their being spared by extinction. They were devastated- though they fared far better than the dinosaurs.
Their long-term success was due to their adaptability in the aftermath, when they exploited the opportunities that presented itself when the dust had settled from the impact. The post-extinction environment provided extraordinary opportunities for snakes. For one, snake food- frogs, lizards, small mammals- must have exploded in the aftermath of the extinction, as there were no dinosaurs to eat them. This post-extinction world was almost devoid of predators, leaving snakes with little competition, and abundant food.
Not only that, the dinosaurs themselves were probably predators of Cretaceous snakes, much like modern raptors and carnivorous mammals- they didn’t just eat the snakes’ lunch, they ate the snakes themselves. The dinosaurs’ removal both as competition, and as predators, left the world wide-open for snakes.
In addition to the sudden explosion in diversity revealed by the DNA, fossils suggests snakes were wildly experimental. Strikingly, snakes moved into aquatic niches. The giant Titanoboa lived in freshwater in South America, and the giant Palaeophis and Gigantophis in marine ecosystems. All three grew to immense sizes- up to around 10 meters in length. They probably filled in for the extinct plesiosaurs and mosasaurs, until whales moved into these niches.
Not just that, but the extinction may have had a profound effect on biogeography. The main constraint on distribution for many animals is probably not climate and environment, but instead, other animals- potential predators and competitors. When an ecosystem is highly diverse, all the available niches are filled, making it difficult for invasive species to become established. In the wake of an extinction, many niches are left open, and few competitors exist. This probably lets species move into new habitats.
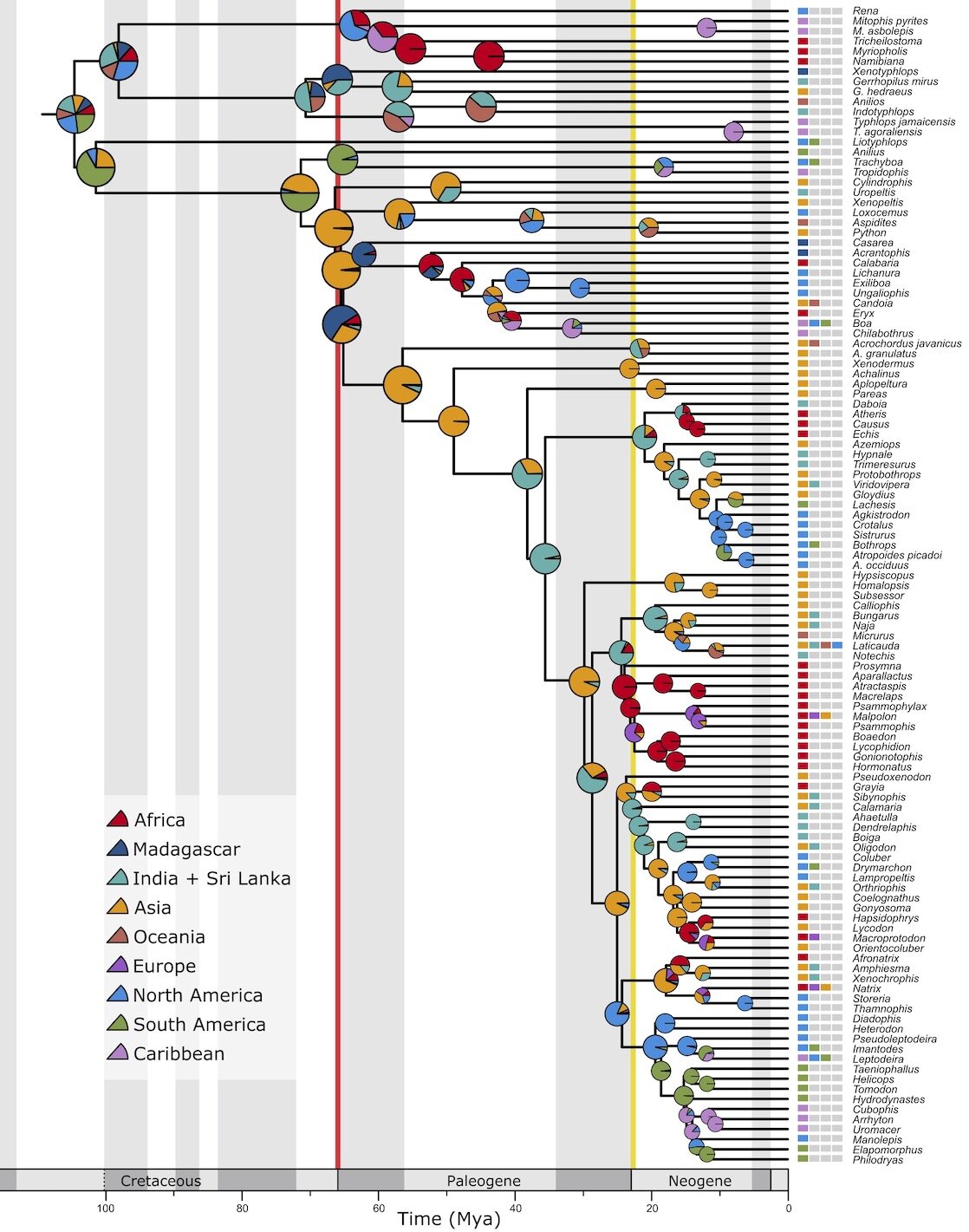
Biogeography of snakes. The orange color of the node near the K-Pg boundary represents a probable Asian range for this ancestor. Snakes are not known from the Cretaceous of Asia- instead they seem to have jumped there after the extinction- perhaps from South America (!)
Strikingly, our analyses suggest a major range expansion of snakes around the time of the extinction- a jump from either Africa or South America, into Asia. While the odds of snakes successfully rafting on vegetation would not have changed between 67 million years ago and 65 million years ago, the massive extinction at 66 million years ago meant that, for species that did raft between continents, they would have found low-diversity ecosystems, with few competitors- and lots of highly edible little mammals and lizards and birds.
Dispersal remained as likely as ever, but the odds of a dispersal becoming established were much higher.
Creative Destruction
Sixty-six million years ago, a huge asteroid struck the Earth. Everything changed, forever.
We know parts of this story well- dinosaurs went extinct (all but a handful of birds). Mammals survived, and flourished. But this is only part of the story. There’s everything else that lived alongside the dinosaurs- snakes, lizards, amphibians, insects, plants- the entire terrestrial ecosystem.
Dinosaurs were extraordinarily diverse- there were hundreds of species. Any event sufficiently extreme to wipe out all dinosaurs, everywhere, must have hit other species as well- the asteroid was not a dinosaur-killing smart missile, but a giant, destructive, asteroid, its destruction global, and not especially discriminating.
Now, slowly, we’ve started to piece the story together, of everything else- moving beyond dinosaurs, and mammals, to other creatures- things like snakes.
Snakes, I’d argue, are interesting not because they’re special, but because they’re not special. They’re an interesting group to study because they’re so diverse, but the pattern they show- severe extinction across the boundary, a major radiation in the aftermath- isn’t special to snakes.
It’s probably the story of terrestrial ecosystems in general.

The Serengeti. Not only the mammals on the Serengeti plain, but the grasses and insects beneath their feet are the result of radiations in the wake of the asteroid impact. It’s arguably the single most important evolutionary event in the past 150 million years- affecting everything.
All the species alive today are the descendants of a few lucky survivors of the asteroid impact. Many, maybe species are the products of these post-extinction radiations. Things that are diverse today, generally speaking, are diverse because they staged radiations after the extinction, not because they weren’t affected. Not just the vertebrates, but everything- insects, plants, fungi.
That grassy meadow? Grasses were rare during the Cretaceous, and grasslands were nonexistent. Grasses became widespread and diverse only after the extinction.
The ants crawling through the grass ? Ants are everywhere today, but were minor players in Cretaceous ecosystems, their dominance is due to post-extinction radiations.
The daisies in that grass hadn’t evolved yet. Neither had the butterflies on that daisy. These things evolved after the dinosaurs went extinct, and in response to the extinction.
Everything alive today descends from species that managed to not only survive the worst disaster in 100 million years, but to compete and thrive in its aftermath. In the Cenozoic, success seems to result not so much from extraordinary resilience in the face of destruction, but from the ability to find opportunity in the wake of disaster.

References
Apesteguía, S. & Zaher, H. A Cretaceous terrestrial snake with robust hindlimbs and a sacrum. Nature 440, 1037-1040 (2006).
Cooper, A. & Penny, D. Mass survival of birds across the Cretaceous-Tertiary boundary: molecular evidence. Science 275, 1113 (1997).
Greene, H. W. Snakes: the Evolution of Mystery in Nature. (University of California Press, 1997).
Head, J. J. et al. Giant boid snake from the Palaeocene neotropics reveals hotter past equatorial temperatures. Nature 457, 715-717 (2009).
Klein, C. G., Longrich, N. R., Ibrahim, N., Zouhri, S. & Martill, D. M. A new basal snake from the mid-Cretaceous of Morocco. Cretaceous Research 72, 134-141 (2017).
Longrich, N. R. The horned dinosaurs Pentaceratops and Kosmoceratops from the upper Campanian of Alberta and implications for dinosaur biogeography. Cretaceous Research 51, 292-308 (2014).
Longrich, N. R., Tokaryk, T. T. & Field, D. Mass extinction of birds at the Cretaceous-Paleogene (K-Pg) boundary. Proceedings of the National Academy of Sciences 108, 15253-15257 (2011).
Longrich, N. R., Bhullar, B.-A. S. & Gauthier, J. A. A transitional snake from the Late Cretaceous period of North America. Nature 488, 205-208 (2012).
Longrich, N. R., Bhullar, B.-A. S. & Gauthier, J. Mass extinction of lizards and snakes at the Cretaceous-Paleogene boundary. Proceedings of the National Academy of Sciences 109, 21396-21401 (2012).
Longrich, N. R., Bhullar, B.-A. S. & Gauthier, J. Mass extinction of lizards and snakes at the Cretaceous-Paleogene boundary. Proceedings of the National Academy of Sciences 109, 21396-21401 (2012).
Longrich, N. R., Scriberas, J. & Wills, M. A. Severe extinction and rapid recovery of mammals across the Cretaceous‐Paleogene boundary, and the effects of rarity on patterns of extinction and recovery. Journal of Evolutionary Biology 29, 1495-1512, doi:DOI: 10.1111/jeb.12882 (2016).
Martill, D. M., Tischlinger, H. & Longrich, N. R. A four-legged snake from the Early Cretaceous of Gondwana. Science 349, 416-419 (2015).
Longrich, N. R., Vinther, J., Pyron, A., Pisani, D. & Gauthier, J. A. Biogeography of worm lizards (Amphisbaenia) driven by end-Cretaceous mass extinction. Proceedings of the Royal Society of London B: Biological Sciences 202, 20143034 (2015).
McCartney, J. A., Roberts, E. M., Tapanila, L. & O'Leary, M. A. Large palaeophiid and nigerophiid snakes from Paleogene Trans-Saharan Seaway deposits of Mali. Acta Palaeontologica Polonica 63, 207-220 (2018).
Prum, R. O. et al. A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing. Nature (2015).
Rio, J. P. & Mannion, P. D. The osteology of the giant snake Gigantophis garstini from the upper Eocene of North Africa and its bearing on the phylogenetic relationships and biogeography of Madtsoiidae. Journal of Vertebrate Paleontology 37, e1347179 (2017).
Vellekoop, J. et al. Rapid short-term cooling following the Chicxulub impact at the Cretaceous–Paleogene boundary. Proceedings of the National Academy of Sciences 111, 7537-7541 (2014).
Wilson, J. A., Mohabey, D. M., Peters, S. E. & Head, J. J. Predation upon hatchling dinosaurs by a new snake from the Late Cretaceous of India. PloS Biology 8, 1-10 (2010).
Notes
Yes, I’m well aware that some people think Tetrapodophis isn’t a snake. My colleagues are entitled to their wrong opinions. Having looked at the anatomy, I stand by the interpretations made in that paper; there’s little if anything I’d change. I don’t claim to be a genius, or infallible; it’s just the evidence is so abundant, so obvious, and so one-sided here. If it looks like a snake, moves like a snake, feeds like a snake, and has all the anatomical features expected for an early snake, it’s a snake. In my opinion the dismissal of Tetrapodophis is perhaps based less on science than on ego and professional jealousy- because there’s simply no scientific argument to think it’s anything other than a snake, and abundant evidence to show that it is. You can find high-resolution figures including all the SI here, and I’m happy to share my photos if anyone’s sincerely curious to know more about the anatomy.
Parviraptor and a number of other snake-like fossils have been identified as Jurassic proto-snakes. While this isn’t impossible- the same idea did occur to me (and that’s also what we got in the Tetrapodophis analysis)- the fossils also show some very primitive features, like unfused parietals, which suggest they’re not closely related to snakes. It’s possible their snake-like teeth and jaws could be convergent on those of snakes. The referral of Parviraptor seems to rest on the assumption that other fossils on the same block of rock don’t belong to Parviraptor. This assumption is based on the fact that (a) they don’t look like snakes, and (b) Parviraptor is a snake- which is sort of circular reasoning.
Reliable identification of fossils is critical here, since a single mistaken identification of a fossil can throw the entire analysis off— a molecular clock analysis is fairly robust against missing data, but it’s extremely sensitive to incorrect data. If an isolated snake vertebra is mistakenly assigned to a modern snake lineage, it would push that lineage and all its sister-lineages far back in time, creating the impression that many branches of the tree are far older than they really are. That means to get accurate analyses, we need to focus on very high quality data, rather than large quantities of data. To deal with this, the analysis focused on using a large number of younger fossils to date splits in the tree; the reasoning being that with living snakes to compare to, we can very confidently identify these fossils to modern families, or even modern genera. It’s a complicated problem, but if you really want the detailed rationale, it’s laid out in the Supplementary Info of the paper.
The other major issue concerns the choice of model. There are different ways to model the ticking of a molecular ‘clock’. A strict clock model assumes everything evolves at the exact same rate. No one really believes this any more. To use an extreme example, would you expect mutations to accumulate at the same rate in the genes of a bacteria and an elephant, when the bacterium has a generation time of hours and a high mutation rate, and the elephant has a generation time of years, and a low mutation rate? Different lineages will tend to accumulate mutations at different rates. Instead, we used a ‘relaxed’ clock model, which allows for things to evolve at different rates.
Critically, we chose a type of relaxed clock that allows for all species to evolve at different rates, but assumes that closely related species tend to evolve at similar rates- that is, a bacterium is likely to evolve at a similar rate to other bacteria, elephants like other elephants.
This is a complicated problem, but there are various reasons to assume that related animals will evolve at rates that are similar to one another. Related species will tend to have similar generation times, which (since mutations build up generation-by-generation) will affect how quickly mutations build up. Related species are likely to have similar mutation rates as well. There are also aspects of the environment which will affect rates of evolution. For example, since species tend to evolve faster in the tropics than temperate regions, a family that inhabits the tropics might be expected to show a faster rate of evolution- vipers in the Rocky Mountains will, all else being equal, evolve more slowly than boas in the Amazon. Or, for example, since species colonizing new habitats tend to evolve rapidly, racer snakes inhabiting the Galapagos might evolve faster than their mainland relatives. Again, it’s a complicated issue, but the reasoning is laid out in the paper.